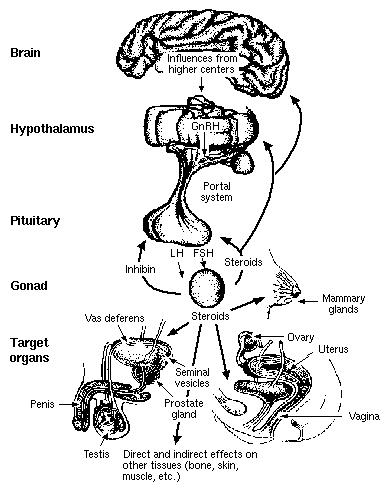
HIPOFIZA SI HORMONII HIPOFIZARI
Luliberina (LHRH, GnRH) Controlul hipotalamic asupra
eliberarii gonadotropinelor hipofizare a fost sugerat de o serie de cercetari
experimentale. Astfel, distrugerea hipotalamusului bazal si a eminentei mediane
provoaca, atat la om, cat si la animal, indiferent de sex, scaderea rapida a
concentratiei plasmatice a LH si FSH, iar stimularea electrica a hipotalamusului
anterior, a zonei nucleului arcuat si a eminentei mediane, determina descarcari
de LH si FSH, care la femeie pot declansa ovulatia. Factorul hipotalamic dotat
cu capacitatea de a stimula eliberarea de gonadotrofine hipofizare a fost
izolat in stare aproape pura de catre colectivele conduse de Guilleminsi,
respectiv, de Schally, i s-a determinat structura biochimica, si apoi a fost
sinetizat .
Diversi autoriau sustinut in
trecut ca hipotalamusul secreta doi hormoni cu actiune stimulanta specifica a
LH si, respectiv, a FSH, hormoni care ar putea fi separati partial din
extractele hipotalamice nepurificate. S-a propus chiar ca acesti doi hormoni
hipotalamici sa fie denumiti LHRH (luteinising hormone releasing hormone) sau
luliberina, si, respectiv, FSHRH (follicle-stimulating hormone releasing
hormone) sau foliberina. Cercetarile recente au dus insa la concluzia
existentei unui singur hormon hipotalamic , care elibereaza ambele
gonadotropine hipofizare, hormon denumit obisnuit LHRH (luliberina), deoarece
efectul asupra eliberarii LH
este mai puternic si mai rapid decat cel de eliberare a
FSH. O alta nomenclatura mult folosita este hormonul eliberator de
gonadotropine (Gonadotropines realeasing hormone - GnRH).
Controlul descarcarilor de LHRH
Controlul descarcarilor de
LHRH este extrem de complex si se realizeaza prin integrarea influentelor
neurale cu cele endocrine.
Cercetarile efctuate la maimute Rhesus au precizat ca la ambele sexe
descarcarile de LHRH in sistemul port hipofizar se fac ritmic, pulsatil, cu
intervale intre pulsuri de 1-3 ore, explicand variatiile ciclice,
caracteristice, ale descarcarilor de LH si FSH. Ritmicitatea descarcarilor de
LHRH reprezinta un element critic pentru mentinerea secretiei gonadotropinelor,
dupa cum reiese din cercetarile in care s-a aratat ca perfuzia continua cu LHRH
mareste initial eliberarea de gonadotropine, dar, daca este continuata mai
multe ore sau zile, raspunsul secretor scade progresiv, adenohipofiza devenind
refractara la actiunea stimulanta a LHRH. Aceste rezultate stau la baza
supresiei secretiei de LH si FSH in pubertate precoce, prin administrarea de
analogi ai LHRH cu durata lunga de actiune. Atat la maimute cat si la femei,
descarcarile pulsatile de LHRH,la intervale de 60-90 minute, stimuleaza
indefinit descarcarile de LH si FSH. Modificarea frecventei descarcarilor
pulsatile de LHRH este urmata la maimute de cresterea sau scaderea selectiva a
concentratiei uneia din gonadotropine, explicand mecanismul prin care un hormon
unic poate modifica raportul LH/FSH.
Cercetarile asupra variatiilor
descarcarilor de LHRH in cursul ciclului menstrual nu au furnizat inca
rezultate concludente, deoarece concentratiile hormonuluihipotalamic in sangele
periferic sunt extrm de mici. Totusi studii preliminare efectuate pe maimute
arata ca secretia de LHRH este crescuta in faza foliculara a ciclului,
concentratiile hormonului in sangele port hipotalamo-hipofizar fiind in aceasta
faza variabile intre valori nedetectabile (sub 10 pg/ml) si atig un maxim de
200 pg/ml . Cercetari efectuate la femeie au evidentiat o crestere importanta a
descarcarii hormonului inaintea ovulatiei si o scadere neta inainte de estrus.
Controlul descarcarilor hipotalamice de LHRH se realizeaza , atat printr-un
mecanism de feed-back negativ, sau pozitiv, declansat de variatiile
concentratiilor plasmatice ale steroizilor sexuali principali (estrogenii la
femei si testosteronul la barbati), cat si prin influente neurale exercitate de
alte formatiuni nervoase.
Componenta endocrina a mecanismului de
control
Secretia gonadotropielor este
supresata la ambele sexe prin administrarea de estrogeni sau de androgeni, si
este stimulata dupa castrare si administrare de droguri antiestrogenice sau
anti androgenice. Efectele de feed-back negativ ale hormonilor steroizi sexuali
se exercita asupra hipotalamusului cat si asupra adenohipofizei, dupa cum
reiese din cercetarile experimentale in care distrugerea hipotalamusului
mediobazal, sectionarea tijei hipofizare sau transplantarea hipofizei au fost
urmate de scaderi profunde ale secretiei de gonadotropine, iar raspunsul
hipersecretor hipofizar la castrare a devenit foarte redus sau chiar absent.
Supresia secretiei de gonadotropine sub actiunea steroizilor sexuali, desi
prezinta diferente intre femei si barbati, implica atat componente hipofizare
cat si neurale. Astfel, administrarea unor doze fiziologice de estrogeni, la
femei cu ciclu normal, este urmata de supresia secretiei bazale de Lh si FSH.
In primele 1-3 zile de administrare responsivitatea hipofizara la doze test de
LHRH este redusa, dovedind ca efectele supresoare ale estrogenilor se exercita,
cel putin partial, la nivel hipofizar. Dupa 3 zile de administrare de
estrogeni, cu toate ca nivelurile bazale ale gonadotropinelor ra man scazute,
hipofiza devine mai responsiva ca in mod normal la LHRH, indicand ca acum,
supresia secretiei gonadotropinelor este datorata inibarii eliberarii
hipotalamice de LHRH.
Administrarea de estrogeni la barbati
este urmata de asemenea de reducerea descarcarilor de LH si FSH, si de supresia
prelungita a responsivitatii hipofizare la LHRH, dovada ca buna parte din
efectul de feed-back negativ al estrognilor la barbati se exercita la nivel
hipofizar, nefiind exclusa insa nici participarea unor efecte inhibitoare
hipotalamice. Administrarea de testosteron la barbati provoaca scaderea
secretiei bayale de gonadotropine, initial neansotita de modificari ale
responsivitatii hipofizare la LHRH, demonstrand implicarea hipotalamusului.
Administrarile prelungite de
estrogeni sau testosteron la ambele sexe determina supresia raspunsului
hipofizar la LHRH, interpretata ca o dovada ca steroizii sexuali principali,
administrati cronic, produc supresia secretiei de gonadotropine prin inhibitie
hipofizara, hipotalamica sau dubla, fara ca raspunsul sa aiba in timp o
evolutie similara. In anumite conditii administrarile de estrogeni la femei
mature sexual pot declansa un efect de feed-back pozitiv, stimuland
descarcarile de gonadotropine prin sensibilizarea raspunsului hipofizar laLHRH.
Acest efect necesitaa, in afara unui hipotalamusnormal, o descarcare acuta de
estrogeni pe fondul unei expuneri cronice la o concentratie bazala scazuta de
estrogeni, conditi realizate de 'valul preovulator' de estrdiol, care
se produce la mijlocul ciclului menstrual. Deoarece la maimute se poate produce
prin administrare de estrogeni cresterea sensibilitatii hipofizare la doze test
de LHRH Si o secretie Crescuta de FSH si LHRH, in conditiile mentinerii
constante a LHRH (animalele avand sectionata tija hipofizara si fiind cu
perfuzie cronica de LHRH), rezulta ca efectul de feed-back pozitiv nu depinde
de cresterea eliberarii hipotalamice de LHRH. Rezultate similare au fost
raportate la femei cu hipogonadism hipogonadotrop prin boli hipotalamice, la
care dupa administrari repetate de LHRH, care maresc progresiv nivelulbazal de
LH si FSH, o injetie cu estrogeni creste suficient nivelul LH ca sa provoace
ovulatie. Aceste constatari au sugerat ca la primate momentul ovulatiei este
determinat de un semnal ovarian, foliculul maturat semnalizand sistemului
hipotalamo-hipofizar oportunitatea declansarii 'valului preovulator'
de gonadotropine.
Componenta neurala a mecanismului de
control
Neuronii hipotalamici care
secreta LHRH primesc variate aferente de la alte etaje ale SNC, cu influente
stimulatoare sau inhibitoare asupra secretiei de gonadotropine, prin aceste
aferente functia sexuala fiind adaptata la modificari;e survenite in mediul
extern sau intern. Miltiple cercetari, efectuate mai ales pe rozatoare -
animale de elctie pentru asemenea studii - au aratat ca la femele expunerea
prelungita la lumina, copulatia si stress-ul, provoaca variatii ale secretiei
de gonadotropine: la femei doar sstress-ul si tulburarile psihice inhiba secretia
gonadotropinica. In producerea acestor modificari secretorii un rol important
au in special sistemul limbic si glanda pimeala .
Sistemul limbic, mai ales amigdala si hipocampul, influenteaza secretia
gonadotropinelor prin intermediul hipotalamusului. Distrugerea sau stimularea
acestor formatiuni nervoase provoaca, in functie de specie, stimularea sau
inhibarea secretiei gonadotropinice, la femei hipocamoul exercitand efecte
inhibitoare, iar amigdala, efecte stimulatoare. Deoarece s-a dovedit ca aceste formatiuni
nervoase poseda receptori pentru estrogeni, se admite ca, in cadrul
mecanismelor de feed-back, exercitate de steroizii sexual asupra secretiei de
gonadotropine, sunt impilcati si acesti centri.
Glanda pineala inhiba eliberarea
de LHRH prin intermediul melatoninei, hormon provenit din serotonina. La multe
specii animale, intre care si la rozatoare, pinealectomia este urmata de
cresterea secretiei de gonadotropine, care la animaleletinere provoaca o
pubertate precoce; injectarea de extracte epifizare are efecte inverse asupra
secretiei gonadotropinice. La mamiferele inferioare epifiza detine un rol
important in cadrul modificarilor secretiei de gonadotropine in functie de
lumina, secretia scazand la intuneric si crescand la lumina, in timp ce la om nu
este inca suficient lamurit rolulepifizei in controlul secretiei de
gonadotropi, desi s-a sratat ca in pinealoame se produce o pubertate precoce.
Descarcarile de LHRH sunt influentate si de unii neurotransmitatori centrali.
Dintre neurotransmitatorii carestimuleaza eliberarea de LHRH mentionam:
acetilcolina, al carei rol este demonstrat de faptul ca atropina in doze mari
blocheaza ovulatia reflexa la iepure, norepinefrina, care injectata in
hipotalamusul mediobazal declanseaza la sobolan descarcarea de LH prin efect
a-adrenergic (a blocantii impiedica raspunsul ovulator la sobolan si iepure),
prostaglandinele, care contribuie la eliberarea gonadotropinelor, actionand la
nivelul ariei preoptice, si nucleului arcuat, si, intr-o masura mai redusa ,
simuland si direct activitatea secretoare a celulelor gonadotrofe
adenohipofizare, histamina, si, probabil, serotonina si dopamina. Dintre
substantele neurotransmitatoare care inhiba eliberarea de LHRH, mentionam:
melatonina epifizara, neurotensina, care exrcita actiunea inhibitoare direct,
endorfinele si encefalinele care actioneaza indirect prin inhibarea actiunii
stimulatoare a dopaminei si prin modularea actiunii inhibitoare a substantei ,
GABA, care actioneaza indirect, inhiband actiunea stimulatoare a dopaminei,
etc. Acesti mediatori ar actiona la toate cele trei nivele ale gonadostatului,
norepinefrina ar exercita un rol stimulator la etapa de integrare a aferentelor
care pot influenta secretia de LHRH, acetilcolina ar interveni la nivelul zonei
neorosecretoare, stimuland secretia de LHRH, iar la nivelul jonctiunii
neurovasculare ar actiona dopamina, favorizand descarcarea hormonului in
capilarele sistemului port hipotalamo-hipofizar, si serotonina, cu efecte
inverse.
B.Axa
hipotalamo-hipofizo-gonadala
Efectele principalilor
neurotransmitatori identificati pana in prezent asupra GnRH implica dopamina si
serotonina. Dopamina stimuleaza eliberarea GnRH, cu toate ca nu este clar daca
aest lucru priveste eliberarea tonica sau ciclica de GnRH. Serotonina exercita
un efect inhibitor atat asupra secretiei ciclice cat si asupra celei tonice de
GnRH.
Norepinefrina poate de asemenea stimula eliberarea de GnRH.
Reglarea axei hipotalamo-hipofizo-gonadala variaza cu varsta si sexul. LH si
FSH sunt prezenti in circulatie de la nastere, iar prin stadiile timpurii ale
pubertatii nivelele de FSH cresc gradual intr-un grad mai mare decat LH. In
timpul acestei perioade raspunsurile FSH la GnRH sunt mai mari deat cele ale
LH, o modalitate de raspuns opus celei observate dupa pubertate, iar
hipotalamusul este excesiv de sensibil la efectele supresive ale steroizilor
gonadali.In perioada prepubertara tarzie (7 pana la 0 ani) secretiile pulsatile
ale LH legate de somn incep, si apare sincronizarea intre pulsurile de LH si
FSH. Aceste pulsuri stimuleaza secretia testosteronului la baieti si de
estradiol la fete, care initiaza caracteristicile clinice ale pubertatii. In
acelasi timp, se observa micsorarea snsibilitatii secretiei hipotalamice de
GnRH ca raspuns la feedback-ul steroid. La femei, dezvoltarea feedback-ului
pozitiv asupra LH si FSH de catre steroizii gonadicigenereaza surge
preovulatorie ciclica a gonadotropinelor, rezultand in instituirea ovulatiilor
ciclice in jurul varstei de 15 ani. La barbatii adulti modelul secretor de LH
este caracterizat de opt pana la zece pulsuri aparand la intervale regulate in
timpul zilei, iar relatia caracteristica adolescentei cu bioritmul somn-veghe
dispare. secretie pulsatila similara a LH si FSH apare la femeile mature,
frecventa si magnitudinea pulsurilor variind cu perioada ciclului menstrual.
Cand foliculii ovarieni dispar la menopauza, secretia hormonilor ovarieni
principali diminua, iar pierderea feedback-ului negativ al acestor hormoni
amplifica eliberarea de FSH si, intr-o masura mai mica de LH. O crestere
similara a si FSH este observata la barbati, intr-a saptea si a opta decada ca
raspuns la scaderea functiei testiculare.
La barbati, stress-ul chirurgical are ca efect o crestere tranzitorie a
nivelului de LH, urmata de o scadere de lunga durata a nivelului de
testosteron. Nici o modificare nu a fost observata la femei, cu toate ca
anovulatia hipotalamica este frecvent observata in timpul stress-ului.
Feromonii si alti factori din mediul inconjurator au fost implicati drept cauza
pentru sincronizarea ciclului menstrual observata la femei ce traiesc impreuna.
Hormonii steroizi regleaza secretie de LH si FSH prin doua mecanisme
principale. Steroizii gonadali regleaza secretia tonica printr-un mecanism de
feedback negativ.Testosteronul pare a fi mult mai puternic decat estrogenii in
aceste efect de feedback negativ, pe cand progesteronul are un efect
intermediar. Inhibina, un peptid produs de celulele granuloase ovariene si de
celulele testiculare Sertoli, are o actiune selectiva in inhibarea eliberarii
de FSH, posibil prin alterarea efectelor GnRH. Eliberarea ciclica a LH si a FSH
este stimulata printr-un efect de feedback pozitiv de catre steroizii ovarieni
in timpul fazei finale a cresterii foliculare, inainte de ovalatie. Cresterea preovulatorie
a LH este precedata de o crestere a nivelurilor estrogenilor circulanti in
prezenta nivelului mic sau descrescator al progesteronului. Efectul pozitiv
actioneaza atat la nivel hipotalamic, cat si la nivel hipofizar, cu toate ca
ultimul pare a fi mai important. GnRH este eliberat Intr-o maniera pulsatila
tonica de hipotalamus aproximativ la fiecare 90 minute, iar acest lodel de
secretia este critic pntru efectele sale pe hipofiza. La o persoana cu secretie
deficitara de GnRH, administrarea pulsatila de Gnrh permite restaurarea
ciclului ovulatoriu normal. In contrast, infuzia constanta a GnRH duce la
fenomenul de 'down-regulation' al receptorilor hipofizari pentru
GnRH, si la o supresie a secretie gonadotropinelor. Acest fenomen a condus la
rezultate importante interapia avand ca scop imbunatatirea sau prevenirea
fertilitatii.
Somatoliberina
(GRH, GHRH)
Numeroase cercetari experimentale si clinice au sugerat controlul hipotalamic
al secretiei hormonului de crestere (GH - growth hormone). Astfel s-a constatat
ca distrugerea experimentala a nucleilor ventromediali hipotalamici produce
scaderea concentratiei plasmatice a GH si oprirea cresterii, corectabila prin
administrarea de extreacte hipotalamice; de asemenea la om tumorile care
distrug hipotalamusul se isotesc de nanism secundar. Stimularea electrica a
nucleilor ventromediali si arcuatidetermina scaderea concentratiei plasmatice a
GH in timpul stimularii, urmata de o crestere ulterioara foarte importanta,
care poate fi blocata prin injectia de somatostatina. Distrugerile hipotalamice
sau grefele de hipofiza in camera anterioara a ochiului sau sub capsula renala
sunt urmate de degranularea celulelor somatotrofe adenohipofizare. Fragmentele
de hipofiza elibereza in vitro substante cu actiune de GH, daca se adauga in
lichidul de incubatie extracte hipotalamice, iar in vivo, injectarea de
extracte hipotalamice crude, sau partial fractionate provoaca eliberare de GH.
Aceste cateva exemple dovedesc existenta unui factor care stimuleaza secretia
si/sau excretia de Gh din adenohiopofiza, factor care a primit denumirea de
hormon eliberator al hormonului de crestere (GHRH - growth hormone releasing
hormone).
Controlul secretiei de GHRH
Controlul secretiei de GHRH se realizeaza de catre mecanisme nervoase si
umorale doar partial cunoscute (fig. ). Diversi stimuli exogeni mentionati
anterior, precum si bioritmul circadian, influenteaza descarcarile de GH prin
intermediul hipotalamusului, respectiv, prin modificarea descarcarilor de GHRH.
Desi secretia hipofizara de GH este controlata de hipotalamus prin 2 hormoni
(GHRG si SRIF), influentele predominante sunt cele stimulatoare, dupa cum
reiese din constatarea ca sectionarea tijei hipofizare sau leziunile
hipotalamusului bazal sunt urmate de inhibitia profunda a descarcarilor bazale
si induse de GH, iar daca se inactiveaza influentele inhibitoare ale
somatostatinei, cresc eliberarea bazala de GH si raspunsurile la stimulii care
obisnuit produc descarcari de GH. S-a sugerat ca inactivarea influentelor somatostatinice
ar explica eliberarea paradoxala de GH dupa injectia de glucoza la unii
pacienti cu gliom al nervului optic (care comprima hipotalamusul anterior),
precum si in diversele forme de encefalopatie metabolica (uremia, coma
hepatica, etc.). Deoarece nici GHRH nici SRIF nu pot fi dozati direct, se
admite ca rezultatul net al oricarui factor care influenteaza secretia de GH
trebuie considerat ca suma efectelor asupra celor 2 hormoni hipotalamici
antagonisti.
Aferentele din hipocamp (legate probabil cu ritmul somn-veghe), precum si cele
de la amigdala bazolaterala, stimuleaza descarcarile de GHRH. Amigdala, fiind o
parte componenta a asa-zisului 'creier visceral', se admite ca ar fi
implicata in descarcarile de GH determinate de emotii Si stresurile fizice.
Neuronii din nucleii ventromediali sunt asociati cu descarcarile de GH in
cadrul mecanismelor reglarii glicemiei, ei continand glucoreceptori care
influenteaza atat secretia de insulina cat si pe cea de GH.
Controlul secretiei de GH de catre hipocamp si amigdala, prin intermediul
nucleilor ventromediali si, respectiv, al descarcarilor de GHRH, este mediat,
cel putin partial, de catre cele 3 sisteme ascendente monoaminergice:
dopaminergic, serotoninergic, si noradrenergic. Astfel s-a aratat ca administrarea
de DOPA, care in creier este convertita in dopamina, produce descarcari de GH
prin stimularea eliberarii de GHRH, atat prin actiune directa asupra
recptorilor dopaminergici, cat si prin stimularea a-receptorilor, dupa
transformare in norepinefrina. descarcarile de GH provocate de somn sunt
mediate in special de fibre serotoninergice si implica descarcari de GHRH; de
altfel, in anumite sindroame hipotalamice, s-a constatat lipsa cresterii
concentratiei plasmatice de GH in timpul somnului. Norepinefrina, prin
stimularea a-receptorilor centrali, este raspunzatoare de descarcarile de GH
induse de hipoglicemie, effort fizic, vasopresina, si cele mai multe stresuri.
Importanta relativa a celor trei sisteme monoaminergice de control difera de la
o specie la alta si de aceea rezulatele experimentale nu pot fi extrapolate la
om. In afara sistemelor monoaminergice, o serie de elte substante hippotalamice
(vasopredina, b-endorfina, peptidul vasoinhibitor (VIP), neurotensina, TRH,
ACTH, a-MSH, GnRH, etc.) stimuleaza de asemenea secretia de GH prin intermediul
GHRH, printr-un sistem de feed-back cu ansa scurta, deoarece aplicate direct pe
hipofiza in vitro sunt inactive. Dar, spre deosebire de ceilalti hormoni tropi
hipofizari, a caror secretie este controlata printr-un mecanism de feed-back de
catre concentratia plasmatica a hormonilor glandelor tinta, secretia de GH este
reglata si de un factor dependent de GH: IGF-1, somatomedina C sau factorul de
sulfatare produs in ficat.. IGF-1 regleaza secretia de GH prin inhibarea
descarcarilor de GHRH din neuronii secretori hipotalamic si stimularea
eliberarii de somatostatina.
Gh insusi, printr-un mecanism de feed-back cu ansa scurta, inhiba eliberarea de
GHRH. Acest mecanism este demonstrat de constatarea ca perfuzia de GH la om
inhiba descarcarile hipofizare de GH induse de hipoglicemia insulinica, iar la
animale eliberarea de GH este inhibata prin implantarea in hipotalamus de
fragmente de tumori hipofizare secretante de GH.
Axa
hipotalamo-hipofizo-somatotropa
Secretia de GH este reglata prin hormoni eliberatori si inhibitori: GRH si SRIF
(somatostatina). GH este secretat intr0o maniera pulsatila cu un nivel bazal la
sau sub nivelulde detectie, peste care se suprapun pulsurile de GH legate de o
ritmicitate neuronala innascuta ce are caefect eliberarea atat a GRH cat si a
SRIF. Majoritatea secretiei pulsatile de GH apare la aproximativ o ora de la
inceputul somnului si este asociata cu fazele de smn 3 si 4. Cu varsta,
secretia de GH se schimba dramatic, atat cantitativ cat si alitativ. Nivelele
extrem de ridicate existente in primele zile de viata scad incepand cu
saptamana a doua de viata. In timpul perioadei pubertare nivele comparabile cu
sau mai mari decat cele de la adult sunt observate, si cresteri ale nivelului
de Gh au loc mult mai frecvent. Dupa a patra decada, are loc o scadere continua
si progresiva in secretia spontana de GH, si raspunsurile la stimulieliberatori
de GH de asemena scad. Reglarea de catre neurotransmitatori a secretiei de GH a
foat extensiv cercetata. Dopamina, norepinefrina (prin receptorii a),
epinefrina, serotonina, GABA, si acetilcolina au fost toate demonstrate a
stimula secretia de GH. Melatonina are atat efect stimulator cat si efect
inhibitor, TRH si CRH exercita efecte inhibitorii (ambele peptide stimuleaza
eliberarea SRIF), iar endorfinele-encefalinele au efecte stimulatorii , toate
mediate In interiorul SNC. Secretia de GH este afectata profund de alimante.
Cresterea nivelului aminoacizilor, descresterea celui al acizilor grasi liberi
si hipoglicemia stimuleaza secretia de GH, pe cand hiperglicemia inhiba
eliberarea de GH. Secretia de GH este crescuta de exercitiu, anxietate, si
stress emotional sau psihic. Multi hormoni afectteza secretia de GH.
Administrarea estrogenilor creste responsivitatea GH, pe cand deficienta sau
excesul de hormoni steroidieni si tiroidieni scad responsivitatea. La barbatii
pubertari sau prepubertari, administrarea androgenilor de asemenea cresc
saspunsul GH. Secretia de GH este dimorfica sexual: famaila au nivele bazale
mai mari si pulsuri mai mici decat au barbatii. Aceste difirinte pot, in parte,
contribui la diferentierea legata de sex in modelele de crestere si in
activitatea enzimatica selectata.
In afara de acesti stimuli 'cu bucla deschisa' exista un sistem de
feedback cu bucla inchisa. GH stimuleaza producerea somatomedinei C (insulin
like growth factor 1 [IGF1] de catre numeroase tesuturi, si atat GH cat si IGF1
exercita efecte de feedback. IGF stimuleaza eliberarea somatostatinei, inhiba
pe aceea a GH, si de asemenea inhiba secretia bazala si stimulata de GRH si
eliberarea din hipofiza a GH. GH stimuleaza secretia SRIF si inhiba expresia
genei ce codifica GH, si secretia GH.
Somatostatina
(SRIH, SRIF)
Somatostatina a fost descoperita de Krulich si colab. (1968), in cursul unor
cercetari in care se incerca izolarea din extractele hipotalamice a hormonului
eliberator de somatotropina. Factorul care inhiba puternic in vitro eliberarea
de GH din hipofizele incubate, a fost de numit hormonul inhibitor al eliberarii
hormonului de crestere (Growth hormone release inhibitng hormone - GHRIH) si
s-a postulat ca secretia GH hipofizar este controlata atat de un factor
stimulator, cat si de un factor inhibitor. Descoperirea a trecut aproape
neobservata, deoarece cei mai multi cercetatori au crezut ca este vorba de un
efect relativ nespecific. dar dupa cativa ani, Brazeau si colab. (1973) au
izolat din extractele hipotalamice un tetradecapeptid, dotat cu puternice
efecte inhibitoare asupra eliberarii GH, pe care l-au denumit SRIH
(Somatotropin release inhibiting hormone). In acelasi an hormonul a fost
sintetizat de mai multe grupe de cercetatori, ceea ce a permis ezxtinderea
cercetarilor ckinice si exoerimentale relative la secretia si actiunile sale.
Controlul secretiei de SRIH
Controlul secretiei de SRIH este inca incomplet cunoscut, dar cercetarile
preliminare sugereza ca principalele structuri in care se gasesc cele mai mari
cantitati de somatostatina (hipotalamusul, tractul digestiv, pancreasul)
sintetizeaza hormonul in functie de necesitatile proprii si sub influenta unor
stimuli diferiti. Astfel, la caine, introducerea in stomac de lipide, glucoza,
sau hidrolizate de cazeina, provoaca descarcari de somatostatina din pancreas,
iar ultimele doua si din antrul gastric. HC introdus in stomac mareste secretia
de somatostatina a pancreasului si a antrului si inhiba secretia fundului
gastric, iar introdus in duoden stimuleaza eliberarea de somatostatina din
pancreas, antru si fundus. Glucoza introdusa in stomac stimuleaza eliberarea de
somatostatina mai puternic decat daca este introdusa in duoden. Perfuzia de CCK
si secretina creste eliberarea de somatostatina din stomac si pancreas, iar GIP
stimuleaza eliberarea hormonului numai din pancreas, dar nu si din stomac. Efect
stimulant asupra descarcarilor de somatostatina mai exercita si glucagonul si
neurotrasmitatorii sistemului simpatic care actioneaza prin b-receptori.
Somatostatina, eliberata din celulele D prezente in mucoasa tractului digestiv
si pancreas, sub actiunea diversilor stimuli, actioneaza predominant local,
paracrin, partial ajunge in lumenul gastricx si duodenal, si in cantitati
minime in sangele circulatiei sistemice. Acetilcolina si stimularea
a-receptorilor adrenergici inhiba eliberarea somatostatinei.
Controlul secretiei hipotalamice de somatostatina este mai putin ckarificat,
dar, prin cercetari in care au fost distruse caile somatostatinergice sau au
fost administrati anticorpi anti-somatostatina, s-a precizat ca secretia de GH
este dominata de factorii stimulatori, iar descarcarile de somatostatina se
realizeaza, similar celor de GHRH, de catre neuronii monoaminergici
(dopaminergici, noradrenergici si serotoninergici), care prin
neurotransmiatorii lor, concomitent cu stimularea eliberarii de GHRH, inhiba
eliberarea somatostatinei. dar secretia de somatostatina este influentata si de
alti factori, unii cu actiune stimulanta (Ca2+, Na+, T3 si T4, cAMP, VIP.
etc.), altii inhibanta (atropina, acetilcolina, GABA, endorfinele, substanta P,
neurotensina, etc.). Mecanismele prin care se exercite efectele stimulante sau
inhibante ale acestor variati factori nu sunt cunoscute. Descarcarile de
somatostatina mai sunt controlate si printr-o ansa periferica a mecanismului de
feed-back, care influenteaza secretia GH, unul din factorii sintetizati de
ficat sub actiunea GH si care mediaza anumite efexte ale GH - somatomedina C
(IGF-1) - stimuleaza eliberarea de somatostatina, concomitent cu inhibarea
eliberarii de GHRH.
