FIZIOLOGIA NEURONULUI
Neuronul - unitate de baza a sistemului nervos
Conform teoriei actuale, neuronul este unitatea genetica, anatomica, fizologica, fiziopatologica, si trofica a sistemului nervos.
a) Unitatea genetica rezida din faptul ca toate celulele din sistemul nervos au originea comuna in celula mama, neuroblastul.
b) Unitatea anatomica a sistemului nervos este neuronul unic, deoarece intre neuronii unui lant neuronal nu exista continuitate(protoplamatica), ci numai contiguitate, realizata prin atingere la nivelul suprafetelor sinaptice.
c) Unitatea fiziologica (functionala) a sistemului nervos, deoarece el este capabil sa receptioneze si sa transmita, in sens aferent si eferent, o excitatie oarecare.
d) Unitatea fiziopatologica a sistemului nervos, fiind capabil sa raspunda singur la efectul unor influente nocive.
e) Unitatea trofica a sistemului nervos deoarece corpul neuronal (pericarionul) asigurp functionalitatea intregului sistem (dendrite si axon).
Structura functionala a neuronului
Neuronul are o strcutura inalt specializata pentru receptionarea si transmiterea informatiei. Din punct de vedere structural, neuronul prezinta un corp celular (soma) si numeroase prelungiri: unele scurte si ramificate, numite dendrite, si o prelungire unica, de obicei mai lunga, ramificata doar in zona terminala - axonul. Din punct de vedere functional, partile componente ale neuronului se grupeaza in trei regiuni sau zone(fig.1).
1. O regiune receptoare, specializata pentru receptionarea si procesarea informatiilor, reprezentata de dendrite si de corpul celular. La nivelul sinapselor, ca zone de comunicare interneuronala, membrana plasmatica a dendritelor si somei reprezinta componenta postsinaptica si este dotata cu receptori specifici pentru neurotransmitatori. Membrana regiunii receptoare poseda putine sau nu poseda deloc canale ionice voltaj-dependente, prin urmare pragul sau de depolarizare este mare si de obicei aici nu se formeaza PA. Excitantul genereaza doar potentiale locale (PPSE sau PPSI) care codifica informatia in amplitudine, direct proportionala cu intensitatea stimulului;
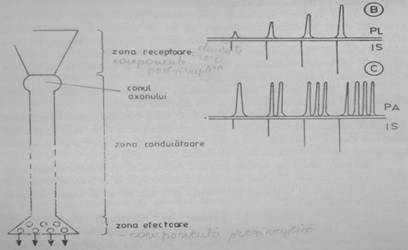

2. O regiune conducatoare, care face legatura intre regiunea receptoare si cea efectoare a neuronului. Este reprezentata de prelungirea axonica, de la locul in care aceasta iese din corpul celular (conul axonic) si pana la arborizatia terminala. Membrana plasmatica a conului axonic prezinta numeroase canale ionice voltaj-dependente. In aceasta zona, prin sumatia PL generate in r egiunea receptoare, ia nastere PA, care apoi se propaga pana la capatul distal al axonului. Se considera ca primul nod Ranvier al axonului ar fi locul de aparitie al PA, in fibrele mielinizate. Dintre toate regiunile functionale ale axonului, conul axonic are cel mai mic prag de depolarizare. Incepand de la conul axonului, informatia este codificicata in frecventa (PA au aceeasi amplitudine, dar frecventa lor este proportionala cu intensitatea stimulului);
3. O regiune efectoare, reprezentata prin butonii terminali ai axonului, care, in cazul comunicarii cu un alt neuron sau cu o celula efectoare joaca rol de componenta presinaptica. Informatia propagata de-a lungul regiunii conducatoare, sub forma de PA, ajunge in regiunea efectoare, unde este recodificata in semnal chimic si transmisa, in aceasta forma, regiunii receptoare a neuronului urmator.
Cele trei regiuni ale neuronului descrise mai sus prezinta particularitati structurale diferite, perfect adaptate functiei.
Corpul celular (soma) si dendritele sunt invelite intr-o membrana plasmatica subtire (neurilem), selectiv permeabila pentru ioni si in consecinta polarizata electric. Canalele ionice sunt preponderent "de scurgere" si ligand-dependente, data fiind functia de componenta postsinaptica.
Nucleul, situat de obicei central, este mare, in ciuda faptului ca neuronul nu se divide. Nucleul este insa direct implicat in intensele sinteze proteice din soma, el detinand informatia genetica ce va fi transmisa prin transcriptie neuriplasmei.
Majoritatea proteinelor, formate de ribozomii liberi si de poliribozomi, raman in corpul celular, in timp ce proteinele sintetizate pe reticulul endoplasmatic rugos (corpusculii Nissl sau corpii tigroizi) sunt exportate spre dendrite si mai ales spre axon. Poliribozomii si reticulul endoplasmatic rugos (RER) sunt caracteristici somei neuronului, axonul nu contine RER si nu poare sintetiza proteine.
Reticulul endoplasmatic neted este implicat in depozitarea
intracelulara a calciului si mentinerea unei concentratii
constante a acestuia, de 10-
In neuroni, aparatul Golgi se gaseste, de asemenea, numai in soma. Ca si in alte tipuri de celule, aceste structuri sunt angajate in glicozilarea terminala a proteinelor sintetizate pe RER si in formarea veziculelor de transport pentru aceste proteine, care sunt eliberate in citoplasma si trimise prin transport axonal spre portiunea terminasa a axonului. Neuronii cu proprietati secretorii (producatori de neurohormoni) din nucleii hipotalamici poseda un aparat Golgi deosebit de dezvoltat.
Transportul proteinelor de la aparatul Golgi depinde de modul de structurare a retelei interne a citoscheletului. Citoscheletul neuronilor este format din microfilamente, neurofilamente si microtubuli.
Microfilamentele sunt formate din actina, o proteina contractila caracteristica celulelor musculare, dar raspandita in multe alte tipuri de celule. Diametrul microfilamentelor este de 4-5 nm; se gasesc mai ales in dendrite.
Neurofilamentele se gasesc atat in dendrite cat si in
axon si sunt formatiunile care confera rigiditatea si
mentin forma neuronului. Nu apar in varfurile de crestere ale
axonilor si in spinele dendritelor, acestea fiind formatiuni mai
dinamice. Neurofilamentele sunt aproximativ de dimensiunea filamentelor
intermediare din alte tipuri de celule )diametrul de
In axoni, transportul anterograd se face cu viteza diferita, in functie de substantele care sunt transportate. Viteza transportului axoplasmic lent este de 1-2 mm/zi. Asa sunt deplasate substante cu rol hranitor, proteine structurale de tipul actinei, neurofilamente si microtubuli. Viteza transportului axoplasmic rapid este de 400 mm/zi; pe aceasta cale ajung in zona sinaptica a axonului organite, vezicule cu mediatori si glicoproteine membranare. Transportul rapid necesita Ca++ si ATP si depinde de rata metabolizarii oxidative a glucozei. Dupa unii autori, ar exista trei tipuri de transport anterograd: lent (1 mm/zi), intermediar si rapid (200-400 mm-zi) iar transportul retrograd ar egala in viteza pe cel anterograd rapid. "Liniile" rapide transporta vezicule cu mediator chimic in sens anterograd si fragmente din membrana veziculelor golite de continut in sens retrograd, spre corpul celular, unde acestea sunt degradate si componentele lor reciclate. Mitocondriile si unele proteine cu rol functional ajung in butonii terminali pe "liniile" cu viteza intermediara, in timp ce proteinele structurale, folosite in cresterea si refacerea prelungirilor, utilizeazp calea de transport lenta.
In dendrite, transportul anterograd se desfasoara cu viteza de 0,4 mm/zi si necesita de asemenea ATP. Prin aceasta modalitate sunt pusi in miscare, pe langa alte substante si ribozomi si ARN, sugerand ca si dendritele sunt capabile de sinteze proteice. De altfel, la baza dendritelor si chiar patrunzand in acestea, pot fi observate concentrari de RER.
Prin transport axoplasmatic retrograd substantele sunt deplasate centripet, dinspre terminatiile prelungirlor spre corpul celular al neuronului. Astfel, substantele ce rezulta din inactivarea si degradarea mediatorilor chimic la nivelul sinapselor sunt recuperate, prin endocitoza, in portiunea terminala a axonului, fiind conduse spre corpul celular si reutilizate pentru noi sinteze.
In cazul unor neuroni, mentinerea integritatii functionale a sinapselor depinde de transportul transneural al unor substante trofice, ca factorul de crestere a nervilor, de-a curmezisul sinapsei. Odata ajuns la soma, prin transport retrograd, factorul de crestere a nervilor activeaza mecanismele sintezei proteice.
Tot antidromic sunt transportate spre corpul celular unele virusuri (herpetic, poliomielitic).
Neurofibrele NU conduc impulsul nervos; acesta se propaga la nivelul neurilemei si axolanei, prin mecanisme pe care le vom studia la subcapitolul despre conductibilitatea neuronului. S-a constatat ca viteza de transport de-a lungul neurofibrilelor este constanta pentru una si aceeasi substanta, indiferent de tipul de fibra nervoasa, in timp ce viteza de conducere a influxului nervos difera in functie de tipul de fibra.
In corpul celular al neuronului se gasesc numeroase mitocondrii, ce furnizeaza ATP-ul necesar intenselor procese de sinteza a proteinelor (inclusiv a mediatorilor chimici). Dar, cea mai mare concentrare a mitocondriilor se afla la nivelul regiunii terminale a axonului, unde nu numa ca produc ATP-ul utilizat ca sursa de energie in procesele asociate transmiterii sinaptice, ci furnizeaza si substrate pentru sinteza unor substante cu rol de neurotransmitatori, ca de exemplu aminoacidul glutamat. Mai mult, mitocondriile contin enzime implicate in degradarea moleculelor de mediatori chimici, ca monoaminoxidaza(MAO), care degradeaza catecolaminele.
Axonul, prelungire unica, lunga, numita de aceea si fibra nervoasa, conduce centrifug influxul nervos (PA) generat la nivelul conului axonic prin sumarea PL. Axonii neuronilor formeaza caile de conducere ascendente si descendente din SNC, precum si nervii periferici. Spre deosebire de dendrite, axonii sunt ramificati numai la capatul distal, unde formeaza arborizatia terminala. Fiecare ramificatie a arborizatiei are un buton terminal, structura implicata in transmiterea sinaptica, mediata chimic, a influxului nervos de la un neuron la altul sau la organele efectoare. Axonii unor neuroni eliberaza mediatorii chimic din proeminente aflate pe traiectul lor numite varicozitati.
Axoplasma este strabatuta pe toata lungimea sa de neurofibrile, care patrund pana in butonul terminal. Axolema este invelita de trei teci: teaca de mielina, teaca lui Schwann si teaca lui Henle.
Teaca lui Henle se afla la exterior si este de natura conjunctiva, asigura nutritia si protectia neuronului. Sub ea se gaseste teaca lui Schwann, formata din celule gliale (celule Schwann), care adera la axolena si se rasucesc in jurul axonului. Teaca de mielina, cu rol de izolator electric, situata in jurul axolemei, este produsa de celulele Schwann si are structura lipoproteica, fiind de fapt o repliere a membranei celulei Schwann, care se rasuceste in jurul axonului. Fibrele nervoase care poseda teaca de mielina bine dezvoltata sunt fibre mielinice, cu viteza mare de conducere a influxului nerovs. La fibrele amielinice, care au si ele teaca Schwann, replierea membranei acestor celule in jurul axonului este redusa si teaca de mielina nu se formeaza. In fibrele amielinice viteza de conducere a influxului nerovs este mai mica decat in cele mielinice.
Doua celule Schwann invecinate nu se ating, intre ele exista un spatiu numit strangulatie (nod) Ranvier, la nivelul caruia mielina lipseste. Strangulatiile Ranvier permit conducerea saltatorie a influxului nervos si formarea colateralelor axonului, cu rol important in stabilirea conexiunilor interneuronale multiple.
Din punct de vedere al numarului de prelungiri, neuronii pot fi unipolari (cu o singura prelungire, axonica, lunga si ramificata - unii neuroni din coretxul cerebelor), pseudo - unipolari (cu o singura prelungire, axonica - neuronii senzitivi din ganglionii spinal), bipolari (cu dendrita si axonul iesind din corpul celular la poli opusi - celulele olfactive, neuronii bipolari din retina) sau multipolari (cu mai multe dendrite si un singur axon - neuronii multipolari din retina, celulele mitrale din bulbii olfactivi, celulele piramidale Betz din cortexul motor). Unii neuroni sunt lipsiti de dendrite (neuronii senzitivi din ganglionii spinali) iar altii nu au axon (celulele orizontale si amacrine din retina).
Din punct de vedere
functional, neuronii pot fi impartiti in trei clase:
neuroni aferenti sau senzitivi, neuronii eferenti sau motori si
neuroni intercalari. Neuronii
aferenti (lat. ad= la; fere= a transporta) duc informatia
de la tesuturile si organele corpului
La capatul distal fata de SNC, neuronii aferenti sunt in legatura cu receptori care, ca raspuns la variatii fizice sau chimice din mediul lor, fac ca in neuron sa se produca semnale electrice. Receptorii pot fi portiuni specializate ale prelungirii neuronale sau celule receptoare separate, aflate in relatie functionala de tip sinaptic cu prelungirea neuronului.
Neuronii aferenti conduc aceste semnale de la periferia organismului spre nevrax. Acesti neuroni sunt atipici, prin faptul ca le lipsesc dendritele si au o singura prelungire, de obicei foarte lunga si mielinizata, considerata a fi un axon. La mica distanta dupa iesirea din corpul celular, axonul se divide; portiunea periferica vine in contact cu receptorul, iar portiunea centrala intra in nevrax, unde stabileste legaturi cu alti neuroni. Corpul celular si portiunea periferica, lunga, a axonului se afla in afara SNC si numai o parte din capatul central patrunde in maduva sau in creier.
Corpul celular si dendritele neuronilor eferenti sunt gazduite in SNC, in timp ce axonii se prelungesc catre periferie, unde stabilesc sinapse cu organele efectoare. Axonii neuronilor aferenti si eferenti, exceptand o scurta portiune, formeaza nervii (sistemul nervos periferic).
Neuronii intercalari se gasesc in intregime in interiorul SNC. Ei reprezinta 99% din totalul neuronilor si au o mare varietate de forme, proprietati fiziologice, mediatori chimici si functii. Pentru fiecare neuron eferent care iese din SNC exista 10 - 20 neuroni aferenti si in jur de 200.000 de neuroni intercalari. Numarul de neuroni intercalari ce se interpun intre anumiti neuroni aferenti si eferenti este diferit, in functie de complexitatea activitatii indeplinite. De exemplu, arcul reflexului rotulian nu cuprinde interneuroni, axonul neuronului aferent (portiunea centrala) se termina pe soma neuronului eferent. Spre deosebire de acesta, reflexele complexe implicate in vorbire sau in procesele de memorie pot cuprinde milioane de neuroni intercalari.
Structura functionala a sinapsei
Desi exista o mare varietate de sinapse din punct de vedere al formei si dimensiunilor componentelor cateva caracteristici de baza sunt comune tuturor acestor formatiuni, sinapsa dintre doi neuroni sau dintre un neuron si o celula efectoare fiind o structura care apartine ambelor celule:
- componenta presinaptica este reprezentata de unul dintre butonii terminali ai axonului neuronului presinaptic (fig. 2). Neurofibrilele, raspunzatoare de transportul orto- si antidromic al substantelor chimice si organitelor celulare, patrund pana in axoplasma butonului terminal, unde se recurbeaza. In jurul neurofibrilelor se afla numeroase mitocondrii ce furnizeaza energia necesara transmiterii sinaptice, precum si vezicule cu mediator chimic (in cazul sinapselor chimice). Veziculele sunt structuri membranare sferice, cu diametrul cuprins intre 300 si 1600 nm, care pot contine unul sau mai multe tipuri de neurotransmitatori. Tehnici speciale de colorare folosite in microscopia electronica au aratat ca membrana presinaptica (portiunea din membrana butonului terminal care este implicata in sinapsa) poseda o retea de filamente care poate conduce veziculele spre situsurile active ale membranei, unde ele pot fuziona cu aceasta;
- componenta postsinaptica este reprezentata de dendrita sau de soma, in unele cazuri de o portiune din axonul neuronului postsinaptic; membrana acestei parti a neuronului, sau membrana unei celule a organului efector pe care se termina butonul terminal, poarta numele de membrana postsinaptica si are o structura foarte diferita de a membranei presinaptice.
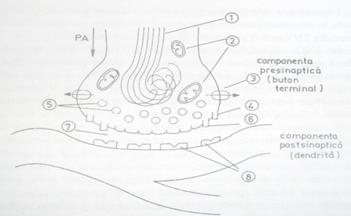
Fig. 2. Structura functionala a sinapsei. 1 - neurofibrile; 2 - mitocondrii: 3 -
pompa de calciu; 4 - canale de calciu; 5 -vezicule cu mediator chimic:
6 - situs activ al membranei presinpatice unde veziculele cu mediator
fuzioneaza cu membrana butonului terminal; 7 - fanta sinaptica;
8 - receptori ai membranei postsinaptice.
fanta sinaptica este un spatiu extracelular, cu o largime de 30-50 nm, care s
fanta sinaptica este un spatiu extracelular cu o largime de 30-50 nm, care separa componenta presinaptica de cea postsinaptica. In cazul sinapselor chimice, acest spatiu este suficient de mare pentru a prezenta rezistenta electrica si a impiedica
trecerea influxului nervos, sub forma PA direct de la o celula la alta. La nivelul fantei sinaptice, trecerea influxului nervos la celula urmatoare este mediata chimic, prin molecule de neuro-transmitator care sunt eliberate din membrana presinaptica si difuzeaza spre membrana postsinaptica.
Deosebirile de structura dintre membrana pre - si postsinaptica (membrana presinaptica nu are receptori conectati la canale ionice ligand-dependente iar compoenta postsinaptica nu contine vezicule cu mediator chimic) duce la o polarizare functionala, adica transmiterea influxului nervos se poate face intr-un singur sens, de la componenta pre la cea postsinaptica.
Importantele diferente functionale dintre sinapsele din SNC somatic, sinapsele neuro-musculare ale musculaturii scheletice si sinapsele neuro-efectoare din SNV rezida in deosebirile de structura functionala.
In SNC, suprafata de membrana neuronala care participa la realizarea unei sinapse este relativ mica, iar depolarizarea produsa in membrana postsinaptica de o sinapsa individuala reprezinta o parte infima din pragul de depolarizare al unui neuron postsinaptic. Din acesta cauza, zeci sau chiar sute de sinapse trebuie sa contribuie la depolarizarea membranei postsinaptice pana la prag, pentru a se putea genera un PA.
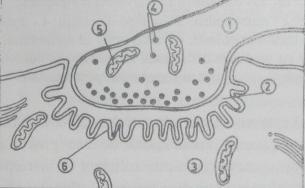
In sinapsele muro-musculare, butonul terminal intra in relatie cu o suprafata mare din membrana fibrei musculare, care la nivelul sinapsei este pliata (fig. 3). Aceasta structura restange pierderile de mediator chimic ca urmare a difuziunii acestuia in fata sinaptica si ofera o arie mai larga de actiune moleculelor de mediator. Fiecare fibra musculara are o singura sinapsa si orice PA care ajunge la butonul terminal produce o depolarizare suficient de mare in sarcolema pentru a atinge pragul si a initia un PA in componenta postsinaptica.
Fig. 3. Sinapsa neuro-musculara (placa
motoare). 1-componenta presinaptica. 2-fanta
sinaptica; 3-compoenenta postsinaptica. 4-vezicule cu mediator
chimic; 5-mitocondrii; 6-membrana postsinaptica (sarcolema)
cutata.
In sistemul nervos vegetativ, neuronii eferenti patrund in organele efectoare si elibereaza molecule de mediator chimic dintr-o serie de dilatatii sau varicozitati. Distanta dintre varicozitati si membrana celulei efectoare este mai mare decat fanta sinaptica din SNC. Ca urmare, mediatorul difuzeaza spre o arie mai larga a membranei postsinaptice. Suprafata de membrana a celulei efectoare care poseda receptori postsinaptici este mai extinsa. Astfel de sinapse mediaza mai incet transmiterea PA, iar raspunsurile efectorului sunt prelungite. Distributia larga a receptorilor face ca membrana postinaptica sa fie sensibila nu numai la mediatorii chimici, ci si la hormoni, care pot fi molecule similare sau chiar identice mediatorilor, aduse de sange (ca adrenalina si noradrenalina, care sunt mediatori ai sinapselor neuroefectoare in SNVSy si la hormonii glandelor medulosuprarenale).
Clasificarea sinapselor poate fi facuta dupa criterii structurale sau functionale.
Din punct de vedere structural, sinapsele se pot realiza intre doi neuroni (neuro-neuronale) sau intre un neuron si o celula a unui organ efector (sinapse neuro-terminale sau neuro-efectoare). Sinapsele neuro-neuronale pot fi axo-dendritice, cand componenta postsinaptica este o dendrita, axo-somatice, cand componenta postinaptica este reprezentata de membrana corpului celular, sau axo-axonice, cand axonul presinaptic se termina pe axolema sau chiar pe un buton terminal al axonului neuronului postsinaptic.
Sub aspect functiona, pot fi clasificate dupa mecanismul de transmitere a influxului nervos, in sinapse chimice, la nivelul carora mesajul electric este recodificat intr-o forma chimica si astfel ajunge la neuronul postsinaptic, si sinapse electrice, prin care PA se transmite neuronului urmator ca atare, fara mediatie chimica.
Dupa natura starii generate la nivelul membranei postsinaptice, sinapsele chimice pot fi excitatoare, cand reactia componentei postsinaptice la mediatorul chimic este o depolarizare urmata de generarea unui nou PA, sau inhibitoare, cand mdoficcarea de potential electric in membrana postsinaptica este o hiperpolarizare si nu se genereaza un nou PA.
Transmiterea sinaptica. Se realizeaza prin doua tipuri de transmitere electrica si chimica.
Functionarea sinapselor elecrice. Sinapsele electrice sunt regiuni functionale in care membranele celor doi neurono sunt foarte apropiate (fig. 4) si legate prin canale membranare de sodiu, caracteristice joctiunilor celulare "gap". Aceste canale permit trecerea libera a curentului electric, astfel ca un PA ajuns la o sinapsa electrica trece in neuronul urmator fara intarziere la nivelul sinapsei. Cu cat suprafata de contact sinaptic intre membrane este mai mare, cu atat schimburile de curent electric intre componentele sinapsei voi fi mai mari. Sinapsele electrice, spre deosebire de cele chimice, permit trecerea influxului nervos in ambele sensuri, neexistand deosebiri structurale majore intre cele doua componente sinaptice.
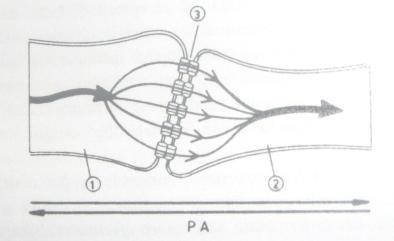
Fig. 4.
Sinapsa electrica; 1 - neuron presinaptic; 2 - neuron
postsinaptic
Sinapsele electrice sunt caracteristice sistemului nervos al nevertebratelor si cea mai importanta functie a lor este sa sincronizeze neuronii, care in mod normal sunt activati deodata. In increngatura vertebratelor, le intalnim in organele electrice ale unor pesti sau intre neuronii care activeaza in mod sincron aripioarele pectorale la pestii zburatori, dar si la vertebratele superioare si chiar la om, in nucleii vestibulari din bulb, unde se cer reactii nervoase foarte rapide, necesare mentinerii sau restabilirii pozitiei corpului in spatiu.
Functionarea sinapselor chimice. Presupune traducerea PA in semnal chimic la nivelul componentei presinaptice, transmiterea in aceasta forma la componenta postsinaptica, urmata de regenerarea PA in neuronul postsinaptic. Eliberarea mediatorului chimic din componenta presinaptica incepe odata ce la membrana butonului terminal ajunge PA condus de-a lungul axonului neuronului presinaptic. Depolarizarea membranei butonului terminal produce deschiderea canalelor voltaj-dependente pentru Ca++ si ionii de calciu patrund din spatiul extracelular in buton, in virtutea gradientului electrochimic. Aceasta determina fuzionarea veziculelor ce contin mediator cu membrana presinaptica, in "zonele active" ale acesteia.
Membrana veziculei si membrana presinaptica se rup la locul de fuzionare si mediatorul chimic este eliberat in fanta sinaptica prin exocitoza. Cantitatea de mediator chimic eliberata este proportionala cu marimea influxului de calciu in butonul terminal. Numarul de molecule de mediator chimic eliberate dintr-o singura vezicula sinaptica poarta numele de cuanta de mediator. Succesiunea evenimentelor din componenta presinaptica care au drept rezultat eliberarea mediatorului chimic sunt ilustrate in fig. 5.

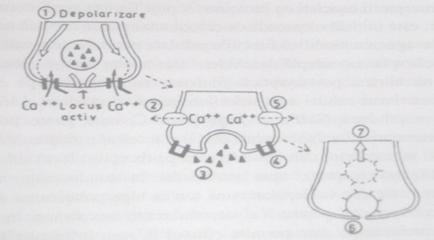
Nu se cunoaste cu precizie mecanismul prin care patrunderea Ca++ in butonul terminal duce la fuzionarea veziculelor cu membrana postsinaptica. O ipoteza considera ca veziculele sunt ancorate de componente ale citoscheletului prin sinapsin, o proteina care inconjura veziculele. Patrunderea calciului in componenta presinaptica ar duce la fosforilarea acestei proteine si deprinderea ei de citoschelet, permitand astfel veziculelor sa fuzioneze cu membrana presinaptica.
Moleculele de mediator chimic difuzate in fanta sinaptica sunt recunoscute de receptorii membranei postsinaptice si se fixeaza (o parte din ele) pe acestia. Receptorii, specifici pentru un anumit mediator, se pot incadra in una din urmatoarele doua categorii:
-receptorii care fac parte din insasi structura canalelor ionice, si
-receptorii care sunt cuplati cu canalul ionic printr-o proteina G si un sistem al celui de-al doilea mesager.
In receptorii asociati cu proteina G, prin fixarea mediatorului chimic pe receptor, este initiata o cascada de reactii enzimatice care da nastere unui al doilea mesager, ce modfica functiile celulare pentru o perioada mai lunga de timp decat o face o simpla deschidere a canalelor ionice. Aceste enzime legate de membrana postsinaptica si mesagerii secundari pe care ele ii produc in interiorul celulei sunt adenilatciclaza ce produce AMPc, guanilatciclaza ce produce GMPc si fosfolipaza C, care produce doi mesageri secundari: diacilglicerolul (DAG) si inozitol trifosatul (IP3).
Cand un mediator chimic se fixeaza pe receptor, formand complexul [MCh-R] (ligand-receptor), apar modificari in conductanta membranei postsinaptice, care duc la depolarizarea sau la hiperpolarizarea acesteia. O crestere a conductantei pentru Na+ depolarizeaza membrana, in timp ce o crestere a conductantei care permite efluxul K+ sau influxul C1- , o hiperpolarizeaza. In anumite cazuri, hiperpolarizarea membranei postsinaptice poate apare si ca urmare a descresterii conductantei pentru Na+, ceea ce reduce influxul de ioni pozitivi.
Intre sosirea unui PA la componenta presinaptica si modificarea de potential in membrana postsinaptica se scurge o perioada de 0,5 - 0,7 ms, cunoscuta sub numele de intarziere sinaptica. Aceasta intarziere este data de timpul necesar pentru patrunderea Ca++ in butonul terminal si fuzionarea veziculelor pline cu mediator cu membrana presinaptica. Timpul necesar mediatorului pentru a difuza in spatiul sinaptic este neglijabil.
Odata deplarizata sau hiperpolarizata, membrana postsinaptica, canalele ionice din aceasta se inactiveaza, inchiderea lor fiind determinata de incetarea actiunii mediatorului chimic. Actiunea mediatorului se opreste cand acesta este:
indepartat de pe receptor prin desfacerea enzimatica a complexului MCh-R];
transformat chimic intr-o substanta inactiva;
transportat, prin mecanisme active, inapoi in butonul terminal sau, in unele cazuri, in celulele gliale invecinate acestuia.
Succesiunea acestor evenimente se desfasoara cu ajutorul unor enzime specifice, in final mediatorul chimic sau substnata rezultata din metabolizarea acestuia fiind recuperata prin endocitoza si transport retrograd de neuronul presinaptic si reutilizata pentru sinteza a noi cantitati de mediator.
Moleculele de mediator chimic care, dupa difuzarea in fanta sinaptica, nu au apucat sa se lege de receptorii postsinaptici vor avea aceeasi soarta ca si moleculele care s-au combinat cu receptorii.
